die geographische lage des städtischen
sees
 Alte Donau, 2015:
Alte Donau, 2015:
Nordbecken (Obere Alte Donau) mit Blick in Richtung Norden über die tiefste Stelle
hin zum Eingang des Strandbades Angelibad. Die tiefste Stelle der Alten Donau, das
Birnerloch, ist 7m tief. Die mittlere Tiefe beträgt 2,5m.
Die Alte Donau (48°14'9.26''N,
16°25'41.6''E) ist
ein flacher See in der Stadt Wien,
der Hauptstadt Österreichs. Der See
ist ein Altarm, der von dem
Donau-Hauptstrom vor etwa 160 Jahren im
Zuge der Flussregulierung abgeschnitten wurde (siehe Karte
Abb.1
in Dokulil et al.
2000 R,
Abb.1
in Donabaum et al.
2004 R).
Dieser Grundwasser-Sickersee
umfasst eine Wassermenge von 3,7 x 106
m3, eine Fläche von 1,5 km2
und hat eine maximale Tiefe von 6,8 m.
Die theoretische Verweilzeit des Wassers betrug üblicherweise etwa 190
Tage, bevor in den letzten Jahren der Wasserspiegel weiter abgesenkt
wurde. Der Altarm ist seit jeher ein beliebtes Naherholungsgebiet. Er
zieht Menschen an, um Boot zu fahren, schwimmen zu gehen oder um
einfach im Grünen entlang des Sees zu spazieren, sich eine kurze
Auszeit in
der Großstadt zu gönnen. Diesem hohen Erholungswert Beachtung
schenkend, hat die Stadt Wien über viele Jahre hier Maßnahmen zur
Verbesserung bzw. später für die Erhaltung der Wasserqualität in der
Alten Donau gesetzt (Dokulil, Donabaum and Teubner 2018 R,
Teubner et al. 2020 R,
2022 R).
Im Jahr 1987, haben Unterwasserrasen von Wasserpflanzen und
Armleuchteralgen (Chara)
fast noch das ganze Wasserbecken der Alten
Donau bedeckt (Abb.4
in Donabaum et al.
2004 R).
Zu jener Zeit setzten
sich die submersen Makrophyten, d.h. die unter Wasser lebenden
Krautpflanzen, gegenüber den im Wasser schwebenden photosynthetischen
Mikroorganismen, wie z.B. den mikroskopisch kleinen Algen, durch (Abb.4
in Dokulil et al.
2000 R)
. Infolge dieser
stabilen Makrophytendominanz
blieb eine Algentrübe im Wasserkörper aus und es war eine hohe Sicht
tief in das klare Wasser hinein gegeben. Die Alte Donau erfuhr jedoch
nachfolgend große Ökosystem-Veränderungen. Der Nährstoffeintrag, vor
allem von Phosphat, erhöhte sich merklich in den Jahren 1988 und 1989
und stieg bis 1994 weiter leicht an. Infolgedessen kam es zu einem
drastischen
Rückgang der Makrophytenbiomasse um bis zu 95%. Mit dem sukzessiven
Verlust und dem letztendlich quasi völligen Ausbleiben der
Wasserpflanzen, dienten die erhöhten Mengen an Phosphat nun alleinig
der Entwicklung der im Wasserkörper schwebenden Algenbiomasse, dem
Phytoplankton. Neben Algen, waren auch Wasserblüten
von Cyanobakterien
Bestandteil dieser Phytoplankton-Massenentwicklungen. Der wesentlichste
Vertreter der Cyanobakterien war hier
Cylindrospermopsis
raciborskii (Abbn.4&5
in Dokulil & Teubner 2000 R)
.
Dieses Cyanobakterium ist allgemein als potentieller Toxinbildner
bekannt und bildete damals bis zu 63% des gesamten Phytoplankton
Biovolumens in der Alten Donau (Seite
1143
in Teubner et al.
2003 R).
 Alte
Donau, 2000:
Alte
Donau, 2000:
Blick auf einen kleinen Teil des südlichen Beckens (Untere Alte Donau). Rechts ist
die Insel "Gänsehäufel" zu sehen. Der Name der Insel bezieht sich auf zwei alluviale
Inseln, die sich vor der Donaufluss-Regulierung und damit vor dem Abtrennen von dem
Hauptflussbett im Jahre 1870 gebildet hatten und die möglicherweise für die
Gänsezucht verwendet worden waren.
 Alte Donau, 2015:
Alte Donau, 2015:
Blick in das kleine Becken "Kaiserwasser", welches mit dem südlichen Becken (Untere
Alte Donau) verbunden ist.
 Alte
Donau, 2015:
Alte
Donau, 2015:
Blick entlang des westlichen Wasserarms des südlichen Beckens (Untere Alte Donau).
 Alte Donau, 2015:
Alte Donau, 2015:
Sicht auf einen kleinen Teil des nördlichen Beckens (Obere Alte Donau) nahe der
U-Bahnstation der U1.
Die interne Seenrestaurierung
der Alten Donau zielte in einem ersten
Schritt auf eine massive Reduktion der Konzentrationen des Phosphats
im
Wasser ab (Abbn.1&2
in Teubner et al.
2003 R),
da dieser Nährstoff allgemein dafür bekannt ist, das
Ausmaß des Wachstums vom Phytoplankton in vielen Binnengewässern zu
bestimmen. In den Jahren nach der erfolgreichen Phosphatreduktion
wurden langfristig andere Maßnahmen der Seenbehandlungen durchgeführt,
und zwar vor allem sogenannte Bio-Manipulationsmaßnahmen.
Ein
wesentliches Augenmerk wurde hierbei auf die Wieder-Ansiedlung bzw.
Neuansiedlung von Makrophyten im See gesetzt (nähere
Information zur
Makrophytenansiedlung bei SYSTEMA und
Teubner et al., 2022 R).
Die Methoden für die Bewertung der Wasserqualität durch Wasserpflanzen
gemäß der Europäischen Wasserrahmenrichtlinie werden im
Detail in Pall & Moser (2009 R)
beschrieben.
Die Erholung von der Eutrophierung während eines frühen Stadiums der Restaurierung wird kurz
in Donabaum et al.
(2004 R) dargestellt.
Die Ökosystemveränderung der nachhaltigen Seerenaturierungen der Alten Donau,
sowohl hinsichtlich abiotischer als auch insbesondere vieler biotischer Aspekte
(von Planktonbakterien und Ciliaten, Phytoplankton und Makrozoobenthos bis hin zu den
Fischen und Wasserpflanzen),
wird für mehr als 20 Jahre in Dokulil, Donabaum und Teubner
(2018 R) aufgezeigt.
Ein in gewissem Umfang, erneutes Auftreten von Makrophyten stellt für sich genommen noch keinen verlässlichen Indikator
für eine erfolgreiche und nachhaltige Gewässersanierung dar. In der Fachliteratur wird daher mehrfach betont,
dass eine reine Wiederbesiedlung durch Wasserpflanzen nicht zwangsläufig mit einer Verbesserung der Wassertransparenz
und somit auch nicht automatisch mit einer Verbesserung des ökologischen Zustandes einhergeht.
Es muss jedoch ausdrücklich hervorgehoben werden, dass diese in der Literatur beschriebenen Beoachtungen
nicht im Widerspruch zu den Untersuchungen in der Alten Donau stehen, da auch hier eine nachhaltige Verbesserung
der Wasserqualität erst nach der flächendeckenden und dichten Ausbreitung der Makrophytenbestände erreicht wurde
(Teubner et al. 2020 R,
2022 R).
Ein lediglich gering ausgeprägtes oder punktuelles Vorkommen von Makrophyten - vergleichbar mit einem sporadischen
Auftreten von Indikatorarten - spiegelt nicht jene ökologischen Rahmenbedingungen wider, die aus stark entwickelten
Makrophytenbeständen (etwa ausgedehnten Unterwasserrasen) oder der Dominanz bestimmter Indikatorarten abgeleitet werden können.
In der Alten Donau zeigt sich jedoch seit den letzten Jahren, etwa ab 2024, dass Makrophytenbestände
zunehmend von watteartigen Fadenalgen überwuchert und dadurch geschwächt werden. Dieses Phänomen,
welches inzwischen auch im Donaudelta weit verbreitet ist (Teubner & Weinbauer, 2025 R), wirft neue Fragen
hinsichtlich der langfristigen Stabilität und Resilienz der Wasserpflanzengesellschaften auf und
könnte den Fortbestand dieser submersen Makrophytenbestände in der Alten Donau nachhaltig beeinträchtigen.
 Alte Donau, 2015:
Alte Donau, 2015:
Südliches Ende nahe dem Pegel Seestern, Untere Alte Donau.
 Alte Donau, 2015:
Alte Donau, 2015:
Nördliches Ende der Oberen Alten Donau mit Blick in Richtung
"Wasserpark".
Die Langzeitstudie der
Seenrestaurierung
der Alten Donau umfasst (Dokulil, Donabaum and Teubner, 2018, R) damit vier
Phasen: (1)
die
nährstoffreiche Situation vor der Sanierung in den Jahren 1993-94, (2)
die nachfolgenden zwei Jahre während der Behandlung mittels Phosphatfällung
(Phosphat Flockung durch RIPLOX-Behandlung Zeitraum 1995-96) sowie weitere drei Jahre für
Massnahmen zur unmittelbaren
Sanierung nach der Phosphatfällung (1997-1999), (3) die nachfolgende Phase der
Wiederansiedlung von Wasserpflanzen über temporäre Wasserspiegelabsenkungen im Jahresverlauf
(2000-2005, nähere Information zur
Makrophytenansiedlung auch in Teubner et al., 2022 R) sowie ab 2006 die Phase eines
stabilen, guten Gewässerzustandes.
Die Vielzahl der abiotischen und biotischen Änderungen bei der Sanierung der Alten Donau
wird über den Zeitraum von mehr als 20 Jahren ausführlich in
Dokulil, Donabaum and Teubner (2018, R) beschrieben.
Warum wurden offensichtlich "lästige Schlingpflanzen " in einem städtischen
Badegewässer angesiedelt?
Spiegelbild einer nachhaltig erfolgreichen Restaurierung eines ehemalig nährstoffreichen
Gewässers ist eine dauerhaft gute (optische)
Klarheit des Wasserkörpers, welche bspw. mittels einer
Secchi-Scheibe gemessen wird
(Sichttiefe ist jene Tiefe bis zu der eine ins Wasser
herabgelassene Secchi-Scheibe unter Wasser noch zu erblicken ist -
je trüber ein Wasserkörper z.B. durch Schwebealgen ist, desto geringer ist die Sichttiefe,
z.B.
Abb.2 in Teubner et al., 2021 R,
R,
R).
Urbane Gewässer eingebettet in grüne Parkanlagen,
die mit ihrem klaren Wasser ein intaktes Ökosystem anzeigen, gelten in Städten dabei als
besonders wertvoll.
Sie werden in vielerlei Hinsicht als gesundheitsförderlich und attraktiv für ein Leben in
urbanen Ballungszentren beschrieben.
Die Erhaltung oder Schaffung solcher sogenannten "blue-green
spaces" ("blaues Gewässer mit grünem Gürtel")
sind somit heute Teil moderner Stadtentwicklung, wie es hier am Beispiel der Alten Donau
gezeigt wird
(Teubner et al. 2020 R).
Während für Badende eine Sichttiefe ab ca. 1.50 m breits akzeptabel erscheint (gewährleitet
einen sogenannten (Durch-)"Blick auf den Seeboden"),
muss für ein nachhaltiges Gedeihen der Wasserpflanzen die Sichttiefe um ein Vielfaches höher
sein
(Teubner et al. 2020 R).
Für die Alte Donau wurde abgeschätzt, dass Unterwasserpflanzen
wie Myriophyllum spicatum
erst bei einer Sichttiefe von mindestens 3,50 m sich langfristig stabil ansiedeln
können
(Abb.4 in Teubner et al. 2020 R,
Abb.3 in Teubner et al. 2021 R,
R).
Der Nachteil von Myriophyllum spicatum ist allerdings,
dass diese Wasserpflanzenart nahe der Wasseroberfläche lange Ausläufer bildet und somit im
Sommer in den Bereichen der
Erholungsnutzung wie Baden und Bootfahren, bspw., unangenehm ist und daher unter Wasser
gemäht werden muss.
Somit stehen Unterwasserrasen gebildet durch Myriophyllum
spicatum in dem Konflikt zwar zum einen Garant zu sein
die Algentrübe des Wasserkörpers zu unterbinden, zu anderen aber für eine Reihe von
Erholungsnutzungen
(sogenannte "Ökosystemleistungen", siehe Abb.4 in Teubner et al., 2021 R,
R) als "Schlingpflanzen"
hinderlich zu sein und somit einen Pflegeaufwand
des Mähens erforderlich zu machen. Ein Ausweg kann z.B. darin gesehen werden, die Sichttiefe
über 3,50 m hinaus zu erhöhen (sommerlich stabil ca. 3,50 bis 4,50 m), um so die Ansiedlung
der am Seeboden flach wachsenden Unterwasserrasen von
Armleuchteralgen zu fördern, wodurch das
kostenaufwendige Mähen nicht mehr notwendig wäre. So ist die Ansiedlung der Armleuchteralgen
auch das eigentliche Ziel der nachhaltigen Sanierung mit stabiler Wassertransparenz in der
Alten Donau -
d.h. die derzeit dichten Myriophyllum-Bestände über eine
natürliche Sukzession durch Armleuchteralgen
ersetzen zu lassen. Solch eine Situation dichter Unterwasserrasen von Armleuchteralgen lag
in den 80iger
Jahren bei einem zugleich hohen Fischbestand
(z.B. Karpfen, siehe Diskussion in Teubner et al. 2022 R)
unter mesotrophen Nährstoffbedingungen vor.
Eine Wassertransparenz von mehr als 3,50 m zum Gedeihen der Wasserpflanzen ist allerdings
weit größer als jene
Sichttiefe von etwa 1,50m, die der Mensch als einen Mindestanspruch an ein Badegewässer
stellt.
Somit tut sich ein Dilemma auf - entweder (1) ein verstärktes Myriophyllum-Wachstum befristet für wenige
weitere Jahre zu akzeptieren und nur unabdinglich notwendige Wasserflächen für das Baden und
Bootsfahren
minimal zu mähen, um eine noch höhere und damit dann auch nachhaltige Wassertransparenz für
das Gedeihen
der Armleuchteralgen voranzutreiben, oder (2) in der Phase einer Wassertransparenz im
Wesentlichen alleinig
gedeihlich für Myriophyllum zu verharren, was sich
zwangsläufig aus der starken Reduktion der Wasserpflanzen
durch Mähen oder sogar gänzlichem Entfernen der Wasserpflanzen sehr frühzeitig in der
"Vegetationsperiode" ergibt
(Unterwassermahd im Spätfrühling bzw. Frühsommer dient der Akzeptanz der Alten Donau als
Freizeitgewässer).
Die Variante 1 bedeutet wohl eher eine gewisse Einschränkung als Badegewässer in Teilen der
Alten Donau für wenige Jahre und
ist daher wohl nur schwer argumentativ aufzubereiten. Die Variante 2 allerdings stimmt
bedenklich wahrscheinlich
die kommenden Jahre unausweichlich recht intensiv Mähen zu müssen, ohne dabei die
Wasserpflanzen zu sehr zu
reduzieren und damit eine unerwünschte Algentrübe wiederum zu riskieren.
Ein Mähen mit Ende der Vegetationsperiode, d.h. z.B. im späten September, ist generell
wünschenswert
um eine Reduktion von Nährstoffen über den Austrag der Wasserpflanzenbiomasse zu bewirken,
ist aber hinsichtlich der Nutzung der Alten Donau als Freizeitgewässers saisonal dann kaum
mehr relevant.
Die Schlüsselrolle der nachhaltigen Sanierung lag in der Alten Donau
ohne Zweifel in der Wiederansiedlung von solchen Wasserpflanzen,
die ganzjährig gänzlich oder überwiegend im Wasser untergetaucht wachsen. Der Grund den
Erfolg der nachhaltigen
Restaurierung der Alten Donau in der Wiederansiedlung der Unterwasserpflanzen zu sehen,
resultiert aus den beiden nachfolgend erklärten Aspekten.
Zum einen ergibt sich mit der vitalen Unterwasserrasen-Entwicklung in der Alten Donau ein
hohes Phosphorspeichervermögen
durch die Wasserpflanzen-Biomasse. Dieses große
Phosphorspeicher-Potential der submersen Makrophyten übersteigt bei weitem die jährliche
Spitzenkonzentration an Gesamtphosphor im Seewasser,
(bezogen auf das Wasservolumen vom gesamten Seebecken), nämlich etwa um das
Zehnfache
(Abb.6 in Teubner et al., 2022 R).
Das einmal in der Wasserpflanzenbiomasse gebundene Nährelement Phosphor ist somit den
Schwebealgen als Nährstoff nicht mehr unmittelbar verfügbar,
wodurch eine Eintrübung des Wasserkörper durch eine Massenentwicklung von Schwebalgen
verhindert wird.
Zum anderen stellen die untergetauchten Makrophyten ein erhebliches Potential eines
zusätzlich verfügbaren Habitates für eine Vielfalt anderer Biota dar.
Dies ist nicht nur für die Fische sondern auch viele kleine Lebewesen relevant, die an den
Wasserpflanzen anheftend leben können (z.B. Makrozoobenthos,
Janecek et al., 2018 R,
Suppl Material data Sheet 4 R in Teubner et al.,
2022 R).
Es konnte für die Alte Donau abgeschätzt werden, dass die
Bio-oberfläche vitaler
Unterwasserrasen um etwa das Zehnfache größer als die gesamten Seeboden-Oberfläche
ist
(Abb.7 in Teubner et al., 2022 R).
Es verlagert sich sozusagen "das Leben im Pelagial", d.h. ein Leben in dem Habitat
Freiwasser, räumlich hin zu einem
"Leben inmitten der Habitat-Architektur der
Unterwasserpflanzen", wodurch wiederum die
Wassertransparenz des offenen Wasserkörpers gefördert wird.
 Alte Donau, 2015:
Alte Donau, 2015:
Die buschigen Wasserpflanzen mit rötlichen Stängeln und Blattrosetten sind selbst
unter Wasser gut in dem klaren Wasser zu erkennen. Diese Wasserpflanze, das Ährige
Tausendblatt (Myriophyllum
spicatum), wächst gut in der Alten Donau und begrenzt somit die
Nährstoffverfügbarkeit gegenüber den Wassertrübe verursachenden Planktonalgen.
Somit gewährleisten die Unterwasserpflanzen die hohe Transparenz des Wassers in der
Alten Donau
(Teubner et al., 2020 R,
2022 R). Ihre dichten
Bestände in der Uferzone dienen darüber hinaus als wertvolle Lebensräume für Fische
und andere Tiere.
 Alte Donau, 2013:
Alte Donau, 2013:
Nach der erfolgreichen Wiederansiedlung von Unterwasserpflanzen im
ganzen See zwecks Verbesserung der Wasserqualität musste der
Ausbreitung dieser Pflanzen nun stellenweise Einhalt geboten
werden.
Das hier gezeigte Unterwasser-Mähboot gewährleistet die Freihaltung der
Hauptfahrrinne und der Uferbereiche mit Badebetrieb von den sogenannten
„Schlingpflanzen“, wie z.B. dem Ährigen Tausendblatt (Myriophyllum spicatum) mit seinen lang-flutenden
Pflanzentrieben.
Können nährstoffreiche Gewässer sich selbst regenerieren?
Es sei an dieser Stelle allgemein erwähnt, dass eine interne Sanierung
ohne gleichzeitige oder im besten Fall sogar voran gestellte Sanierung des
Einzugsgebietes von vorn herein nicht funktionieren kann, weil die
Ursache der Nährstoffbelastung nicht abgestellt wird. Dieser Frage der
Verhinderung des Nährstoffeintrages in die Alte Donau wurde im Vorfeld
der Sanierung nachgegangen. Eine Reduzierung der Nährstoffbelastung im
See wird am kostengünstigsten, wenn ein unabdingbares, nachhaltiges
Management im Einzugsgebiet des Sees erfolgreich durchgeführt wird,
sodass sich zusätzlich teure, seen-interne Behandlungen, wie sie hier
nachfolgend für die Riplox Technik beschrieben werden, erübrigen
können. Es muss hierbei gesagt werden, dass ein See nach Abstellen der
Ursache, d.h. mit der Verhinderung eines fortlaufend, neuen
Nährstoffeintrages, sich selbst über Jahre hinweg wieder "reinigt",
d.h. sich hin zu einem
Gewässer entwickelt, welches seinem ursprünglichen oder natürlichen
„Referenz-Zustand“ entspricht. Diese kostengünstige „allein-externe“
Seensanierung hat nur einen erheblichen Nachteil: das „Sichtbarwerden“
des Sanierungserfolges
wird einige Jahre auf sich
warten lassen. Nach dem Passieren des Wendepunktes aufgrund einer
erfolgreichen Restaurierung des Einzugsgebietes von einem See, würde
das Ökosystem über viele Jahre dem Trend der weiter abnehmenden
Verfügbarkeit von Nährstoffen folgen. Die Zeit, die benötigt wird, um
den ursprünglichen Referenz-Zustand eines Sees wieder zu erreichen,
wird natürlich je nach dem Grad der Verschmutzung, der Größe des Sees
und der Verweilzeit
des Wassers usw. variieren, kann aber nach etwa 4-8 (bzw. 12)
Jahren erwartet werden, was in etwa einem Zeitraum von ein bis zwei
(bzw. drei) Legislaturperioden in vielen Ländern entspricht.
Die Reaktion eines Ökosystems auf sich ändernde Nährstoffbedingungen,
d.h. die Zunahme von Nährstoffen (Eutrophierung)
auf der einen Seite
und die nach einer Sanierung einsetzende Reduktion von Nährstoffen auf
der anderen Seite (Re-Oligotrophierung oder besser Oligotrophierung
genannt), geschehen nicht „über Nacht“, sondern verlaufen über viele
saisonale Zyklen, über Jahre hinweg. Die Ökosysteme reagieren nicht
unmittelbar, sondern verzögert auf Umweltveränderungen
(Ökosystem-Hysterese).
Die natürlich verzögerte Reaktion des
Phytoplanktons auf die zunehmende Nährstoffverarmung
während einer Oligotrophierungsperiode, wird auf dieser Website für den
tiefen alpinen
Mondsee S
beschrieben (siehe auch Seite
2 und Abb.4
in Dokulil & Teubner
2005 R).
Unabhängig von der Art der
Restaurierung eines Sees – ob sie allein über die Sanierung im
Einzugsgebiet läuft oder darüber hinaus durch see-interne
Sanierungsmethoden unterstützt wird, die Informationen über den
Fortschritt einer Sanierung oder Oligotrophierung eines Gewässers kann
nur über
begleitende wissenschaftliche Untersuchungen in Erfahrung
gebracht werden. Mit dem Verstehen der individuellen Prozesse und dem
Erkennen von Tendenzen oder Trends in einem sich ändernden Ökosystem,
können dann
zielgerichtet weitere Sanierungsmaßnahmen abgeleitet und durchgeführt
werden, sofern
sich dies dann als sinnvoll und notwendig ergibt.
 Alte Donau, 2015:
Alte Donau, 2015:
Die Süsswasserqualle (Craspedacusta sowerbii) kommt
nur in manchen Jahren und
nur kurzweilig während heisser Sommerperioden in der Alten Donau vor R. Wenn, so wie auf dem
Foto gezeigt, nur wenige Individuen auftreten, können diese kleinen Medusen im
Wasser leicht übersehen werden.
 Alte Donau, 2015:
Alte Donau, 2015:
Zwei Süßwasserquallen (Craspedacusta sowerbii) und
ein loser Sproß vom Ährigen Tausendblatt (Myriophyllum
spicatum) mit einer Schüssel aus dem Wasser geschöpft.
 Alte Donau, 2015:
Alte Donau, 2015:
Grössenvergleich zwischen der Süsswasserqualle (Craspedacusta sowerbii)
R und der Wasserpflanze
(Myriophyllum
spicatum).
 Alte Donau, 2015:
Alte Donau, 2015:
Eine kleine Süsswasserqualle (Craspedacusta sowerbii)
in der Hand haltend.
Eine Beschreibung dieser Süßwassermeduse aus der Sicht von Tauchern ist z.B. in
Mathä (2009) gegeben; eine Sicht auf diese Art als Neobiota, ihre weite Verbreitung
im Süßwasser sowie ihre Phylogenie sind in Fritz et al. (2007) spezifiziert (siehe
Literaturliste unten).
the implementation of the RIPLOX-method
to reduce phosphate availability for algal growth by phosphate
flocculation in old danube
The Riplox-method
(Ripl 1976, R)
was chosen as the main internal restoration method to
reduce phosphate availability for phytoplankton growth. The method acronym Riplox is
according to the family name of Prof. Wilhelm RIPL, who
described the
lake restoration technique of OXidizing
the sediment surface to prevent internal nutrient loading by the
phosphate release from
anaerobic
sediments in heavily eutrophied lakes. Due to
the costs for in-lake restoration, this technique is usually applied
to smaller, rather than to larger water basins.
In Old Danube, the Riplox-restoration method was implemented
twice, in the spring of 1995 and
1996 (scheme of RIPLOX-treatment in Fig.6
in Dokulil et al.
2000 R).
Two steps were carried out on each occasion: First, FeCl3
buffered with limestone, was added to the water surface to
remove phosphorus and suspended material by chemical and mechanical
flocculation. After this, Ca(NO3)2
was added to the sediments to enhance nitrate oxidation. The mode of
RIPLOX-method is the following: With enhanced nitrate
oxidation, nitrate is then reduced to elemental nitrogen by anaerobic
denitrification. Organic mud is hence oxidized to carbon-dioxide and
water. Consequently, the oxygen deficit that is caused by heterotrophic
metabolism is lowered. Thus, prolonged anoxic conditions at the
sediment surface which is common in a nutrient-rich lake can be
prevented. The
internal phosphate loading by release from the sediment is thus
avoided. This initial treatment in Old Danube indeed shifted the
pelagic community, i.e. the micro-biota drifting in the free water
zone, from nutrient-rich conditions in 1994 to a sustained lower
nutrient status in 1995/1996 as described by the decrease of phosphorus
concentration and the response by the microbial community in detail
below.
 Alte Donau, 2015:
Alte Donau, 2015:
Im Zuge der Seensanierung wurde an vielen Stellen an der Alten Donau das Seeufer neu
gestaltet. Die hier gezeigte Neubepflanzung der Litoralzone im Kaiserwasser gleicht
der natürlichen Vegetationsabfolge an ökologisch intakten Seeufern: ein Schilfgürtel
(Phragmites), der landwärts gerichtet ist, wird
wasserwärts von einem Rohrkolbenbestand (Typha)
abgelöst und geht im flachen Uferbereich zum offenen See hin in eine
Schwimmblattzone über.
 Alte Donau, 2015:
Alte Donau, 2015:
Der Gemeine Wasserschlauch (Utricularia vulgaris)
befindet sich inmitten der im linken Bild gezeigten Schwimmblattzone. Die kleinen
gelben Blüten dieser eher seltenen Wasserpflanze sind nur aus der Nähe zwischen dem
Tausendblatt (Myriophyllum spicatum) deutlich
erkennbar.
 Alte Donau, 2015:
Alte Donau, 2015:
Mit der ökologischen Rekonstruktion der Uferzone am Ostufer der Unteren Alten Donau
(östlicher Wasserarm) wurden wiederum Lebensräume geschaffen, die von vielen Tiere
angenommen wurden. Ein Frosch, zum Beispiel, ist auf der linken Seite des Fotos nahe
den Halmen vom Schilf (Phragmites australis)zu
sehen.
 Alte Donau, 2015:
Alte Donau, 2015:
Das Foto wurde an der gleichen Stelle wie das linkseitige Foto aufgenommen und zeigt
einen Frosch (Rana esculenta) im
Detail.
reduction of total phosphorus: ecosystem
assessment rather by concentrations of total
phosphorus than of phosphate
Phosphorus is known as
the nutrient
element mainly responsible for limiting the growth of algae and
cyanobacteria in many
freshwaters and also some marine systems. Phosphate
dissolved in the water is the only chemical phosphorus species that can
be utilized immediately by photosynthetic organisms as a nutrient
source. A further phosphorus fraction, the dissolved organic phosphorus
can only be utilized via extracellular enzymes released by bacteria and
primary producers. The fraction of bound phosphorus in the living
microorganism cells, i.e. the particulate organic phosphorus, is a
suitable nutrient source for algal growth NOT at all. For these reasons
dissolved inorganic phosphate indeed is used as the nutrient source for
phosphorus uptake
experiments. Such empirical phosphate additions are used
to conduct
algal
culture experiments in the laboratory or bioassay
fertilizing studies with phytoplankton in a lake.
These
experiments may aim at measuring the short-term
course of phosphorus acquisition of algae in the lab, or
the bioassay outgrow
experiments in a longer run over days identifying
potential algal species shift among phytoplankton in the lake.
 Alte Donau, 2000:
Alte Donau, 2000:
Dieses Foto, das im Winter in den Jahren nach der chemischen Seenrestaurierung
(RIPLOX-Phosphatfällung, Ripl 1976)
aufgenommen wurde, verweist auf eine hohe Transparenz des Wassers (Teubner et al.,
2020R,
2022 R). Die
winterliche Sichttiefe des Wassers kann jedoch nicht den trophischen
Zustand eines Ökosystems anzeigen. Die geringe Wassertrübe im Winter
muss nämlich nicht auf eine geringe Verfügbarkeit von Nährstoffen und
somit auch nicht auf ein nährstoffbegrenztes Wachstum von Mikroorganismen im
Wasser (Phytoplankton) verweisen. Der Grund des ungetrübten
Wasserkörpers kann einfach die niedrige Temperatur und kurze Tageslänge
sein, wodurch sich viele Phytoplanktonarten trotz eventuell gutem
Nährstoffpotenzial nicht entwickeln können. Eine massive
Biomasseentwicklung ergibt sich in der gemäßigten Zone in der Regel nur
während der Vegetationsperiode. Zwei oder sogar drei Spitzen der
Biomasseentwicklung zeigen sich im Jahresverlauf: zum Ende vom
Frühjahr, im Hochsommer und manchmal nochmals im frühen
Herbst.Phosphate,
however, appears not to be an
accurate indicator of phosphorus resources
in a lake or river
ecosystem. The analytical detection limit for phosphate, measured as soluble reactive phosphorus
(SRP), depends on the method and is usually about 60-140 nano mol L-1.
In the case where the phosphate concentration in lake water is below
the analytical detection limit, it does not necessarily mean that ‘no’
phosphate is
available for growth and consequently ‘no’ algal biomass may increase.
This evidence comes from field studies. It can be shown that during
periods when phosphate in the epilimnetic lake sample is below the
analytical detection limit for days to weeks, the biomass of algae and
cyanobacteria may even increase at the same time. One reason is that primary producers are able to utilize
phosphate within minutes even below the concentration
range of the analytical detection limit (Teubner et al., in
preparation). A second reason can be seen in
the patchy environment as larger phosphate concentrations are indeed
available but only at very small dimensions of a few millimeters as by
point sources provided by zooplankton excretion (Teubner et al., in
preparation). Such patchy
ephemeral phosphate sources
also seem to be utilized by algae (see page
Preface S
). A third reason might be seen in
the
particular short turnover
time for phosphorus ranging from hours to days. Bound
phosphorus in biota or dissolved organic forms is after short microbial
cycles again available as a dissolved form of phosphate and can
be rapidly utilized. A last reason why phosphate concentration
measures in a
lake or river are often ‘zero’ even at periods of phytoplankton biomass
increase is that regular sampling is usually done during the daytime.
Phosphate
concentrations often varies with the day-night
cycle and are frequently, for some ecological reasons,
considerably higher during the night than daytime (Teubner et al., in
preparation). When phosphate is
limiting the growth of phytoplankton in a lake or river, the night-time
released phosphate can be day-time rapidly incorporated by biota.
Therefore, after enhanced concentrations during the night, the
phosphate can be again within minutes to few hours below the analytical
detection limit at day-time (Teubner et al., in preparation). To meet
these
four concerns, the total
phosphorus concentration
(TP) is usually measured to assess the nutrient state of an aquatic
ecosystem (examples for the Old Danube: Fig.4
in Dokulil et al.
2000 R,
Fig.5
in Donabaum et al.
2004 R,
examples for other lakes on this website see Mondsee S, Traunsee S
and
Taihu S).
In the case study of Old Danube restoration, the concentration of total
phosphorus (TP)
decreased drastically from 2.4 in 1994 (before treatment) to 0.6 µmol L-1
in 1996 (after treatment by RIPLOX-phosphate flocculation) in the water
body. According to ecosystem nutrient-state phytoplankton-response
curves, where concentrations of total phosphorus are plotted versus
concentrations of chlorophyll-a (this pigment that is used as the rough
estimator for biomass of primary producers), Old Danube changed
from a hypertrophic
to a moderately eutrophic ecosystem.
microbial community response to reduced
phosphate availability:
shift toward phosphorus-rich biota under
phosphorus-deficient conditions
 Donau, 2011:
Donau, 2011:
Entnahme einer Phytoplanktonprobe im Frühjahr bei einem praktischen
Algenkurs an der Universität Wien. Die Artenzusammensetzung wird sowohl
anhand einer Lebendprobe als auch an einer fixierten Probe unter dem
Mikroskop später im Labor bestimmt werden.
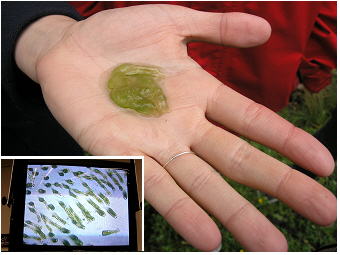 Donau, 2011:
Donau, 2011:
Dieses „grüne Gelee“ wird im Frühjahr an der flachen Uferzone des
Schilfgürtels relativ häufig gefunden. Trotz der grünen Farbe, ist es
in erster Linie keine Alge sondern ein Tier, Ophrydium
versatile.
Das
eingefügte Foto zeigt ein Detail der in der Kolonie lebenden Zellen
unter dem Mikroskop: die einzelligen Tiere, Ciliaten, kontrahieren und
expandieren in periodisch kurzen Zeitabständen. Die grüne Farbe stammt
von
endosymbiontischen grünen Algenzellen, Zoochlorellen, die innerhalb
dieser
tierischen Einzeller leben. Diese Art ist in Seen mit geringfügigem bis
moderatem Nährstoffgehalt weit verbreitet. Sie kann aufgrund der
gelbgrünen Farbe leicht von den dunkelgrün-bräunlichen
„Gallert-Kolonien“
von Nostoc pruniforme
(„Seepflaume“) an den Ufern anderer Seen unterschieden werden.
The ecosystem nutrient status can be
assessed in more detail by the allocation pattern and stoichiometric
proportion among fractions of a
source, as opposed to
the total phosphorus vs chlorophyll-a relationship. Such assessment by
triple ratios is described for the case study Old Danube. Here the
effects
of
reduced total size of phosphorus concentrations from 2.4
to 0.6 mmol L-1 on the plankton community in Old
Danube were evaluated from two triple
ratios. The relative proportions of dissolved reactive
phosphorus (DRP), dissolved non-reactive
phosphorus (DOP: mainly dissolved organic P, and some inorganic
polyphosphates) and particulate organic phosphorus (POP; mainly the
phosphorus incorporated by micro-biota drifting in the water) refers to
the three fractions of total
phosphorus. The partitioning
among the fractions of total
phosphorus is displayed by the DRP:DOP:POP
triple ratio (Fig.4a-d
in Teubner et al.
2003 R).
Secondly, the stoichiometry
among three main nutrient
elements incorporated by biota, refers to the particulate
organic
carbon (POC), nitrogen (PON), and POP, and is displayed by the POC:PON:POP triple ratio (Fig.5e
in Teubner et al.
2003 R)
.
In both triple ratios, POP was used as a key component to indicate
shifts between phosphorus fractions and between nutrient elements of
plankton biota.
At reduced phosphorus availability due to RIPLOX-treatment,
the planktonic assemblage accumulated 14% more phosphorus in their
cells than under phosphor-rich water conditions before treatment. This enhanced phosphorus accumulation by
biota was achieved in two stages. In the first year, it
was
mainly at the expense of DOP and in the second year by DRP (Fig.4b-c
in Teubner et al.
2003 R).
The
evidence indicates that this increase of %POP of TP reflects the
adaptive
phosphorus utilization of the organisms under low phosphorus supply,
substantiated by significant relationships between phosphorus fractions
and changes within the community. A higher %POP of TP was associated
with higher specific enzyme
activity (alkaline phosphatase) and with tighter
coupling between producers and
consumers respectively — that is, driven from both the
nutrient-producer and the
producer-consumer interface (Fig.4e-f
and Fig.5a-d
in Teubner et al.
2003 R).
A lowered %DOP of total
phosphorus was significantly related to smaller overall loss rates for
the carbon pool of bacteria and algae, whereas a lower %DRP of total
phosphorus was correlated with the decrease of the biomass of most
organism groups.
Stoichiometric
shiftstoward a phosphorus-rich assemblage under low TP
have been
justified by compositional shifts of the plankton community, namely
from nitrogen-rich cyanobacteria to phosphorus-rich eukaryotic algae
and bacteria in Old Danube (Fig.5e
in Teubner et al.
2003 R)
. The stoichiometric shift toward the
limiting element seems to be a common phenomenon of individual
adaptation of producer organisms. Studies from other shallow lakes show
that
nitrogen-rich
plankton, i.e. mainly composed by cyanobacteria, dominate usually under
nitrogen-limitation (Figs.1&2
in Teubner et al.
1999 R,
Fig.4a
in Teubner & Dokulil
2000 R).
In turn phosphorus-rich
organisms, i.e. mainly
diatoms and other eukaryotic algae, dominate under phosphorus limitation
(Fig.5 f
in Teubner et al.
2003 R).
The elemental composition of individual cells of bacteria,
cyanobacteria and algae can vary strongly in contrast to consumers
keeping their elemental composition within a limited range, even if the
elemental composition of ingested food is very different from their
body tissue.
In conclusion, the shifts toward POP in both triple ratios
strongly indicated that the plankton
community acted as a sink for phosphorus under reduced total pool size
in Old Danube (abstract
& page 1148
in Teubner et al.
2003 R).
 >
>